
BLASCO Ruth
- Zooarchaeology and Taphonomy, Institut Català de Paleoecologia Humana i Evolució Social (IPHES-CERCA), Tarragona, Spain
- Ancient Palaeolithic, Europe, Middle Palaeolithic, Osseous industry, Spatial analysis, Symbolic behaviours, Taphonomy, Upper Palaeolithic, Zooarchaeology
- recommender, manager
Recommendations: 4
Reviews: 0
Recommendations: 4
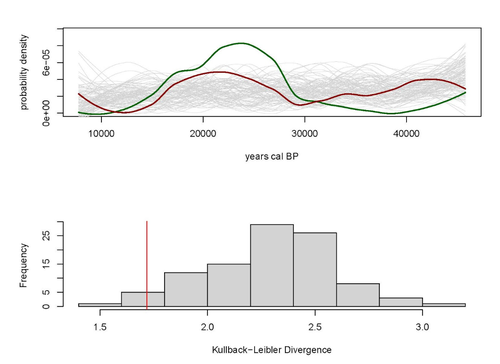
A note on predator-prey dynamics in radiocarbon datasets
A new approach to Predator-prey dynamics
Recommended by Ruth Blasco based on reviews by Jesús Rodríguez, Miriam Belmaker and 1 anonymous reviewerVarious biological systems have been subjected to mathematical modelling to enhance our understanding of the intricate interactions among different species. Among these models, the predator-prey model holds a significant position. Its relevance stems not only from its application in biology, where it largely governs the coexistence of diverse species in open ecosystems, but also from its utility in other domains.
Predator-prey dynamics have long been a focal point in population ecology, yet access to real-world data is confined to relatively brief periods, typically less than a century. Studying predator-prey dynamics over extended periods presents challenges due to the limited availability of population data spanning more than a century. The most extensive dataset is the hare-lynx records from the Hudson Bay Company, documenting a century of fur trade [1]. However, other records are considerably shorter, usually spanning decades [2,3]. This constraint hampers our capacity to investigate predator-prey interactions over centennial or millennial scales.
Marom and Wolkowski [4] propose here that leveraging regional radiocarbon databases offers a solution to this challenge, enabling the reconstruction of predator-prey population dynamics over extensive timeframes. To substantiate this proposition, they draw upon examples from Pleistocene Beringia and the Holocene Judean Desert. This approach is highly relevant and might provide insight into ecological processes occurring at a time scale beyond the limits of current ecological datasets.
The methodological approach employed in this article proposes that the summed probability distribution (SPD) of predator radiocarbon dates, which reflects changes in population size, will demonstrate either more or less variation than anticipated from random sampling in a homogeneous distribution spanning the same timeframe. A deviation from randomness would imply a covariation between predator and prey populations. This basic hypothesis makes no assumptions about the frequency, mechanism, or cause of predator-prey interactions, as it is assumed that such aspects cannot be adequately tested with the available data. If validated, this hypothesis would offer initial support for the idea that long-term regional radiocarbon data contain signals of predator-prey interactions. This approach could justify the construction of larger datasets to facilitate a more comprehensive exploration of these signal structures.
References
[1] Elton, C. and Nicholson, M., 1942. The Ten-Year Cycle in Numbers of the Lynx in Canada. J. Anim. Ecol. 11, 215–244.
[2] Gilg, O., Sittler, B. and Hanski, I., 2009. Climate change and cyclic predator-prey population dynamics in the high Arctic. Glob. Chang. Biol. 15, 2634–2652. https://doi.org/10.1111/j.1365-2486.2009.01927.x
[3] Vucetich, J.A., Hebblewhite, M., Smith, D.W. and Peterson, R.O., 2011. Predicting prey population dynamics from kill rate, predation rate and predator-prey ratios in three wolf-ungulate systems. J. Anim. Ecol. 80, 1236–1245. https://doi.org/10.1111/j.1365-2656.2011.01855.x
[4] Marom, N. and Wolkowski, U. (2024). A note on predator-prey dynamics in radiocarbon datasets, BioRxiv, 566733, ver. 4 peer-reviewed and recommended by Peer Community in Archaeology. https://doi.org/10.1101/2023.11.12.566733

Zooarchaeological investigation of the Hoabinhian exploitation of reptiles and amphibians in Thailand and Cambodia with a focus on the Yellow-headed tortoise (Indotestudo elongata (Blyth, 1854))
A zooarchaeological perspective on testudine bones from Hoabinhian hunter-gatherer archaeological assemblages in Southeast Asia
Recommended by Ruth Blasco based on reviews by Noel Amano and Iratxe BonetaThe study of the evolution of the human diet has been a central theme in numerous archaeological and paleoanthropological investigations. By reconstructing diets, researchers gain deeper insights into how humans adapted to their environments. The analysis of animal bones plays a crucial role in extracting dietary information. Most studies involving ancient diets rely heavily on zooarchaeological examinations, which, due to their extensive history, have amassed a wealth of data.
During the Pleistocene–Holocene periods, testudine bones have been commonly found in a multitude of sites. The use of turtles and tortoises as food sources appears to stretch back to the Early Pleistocene [1-4]. More importantly, these small animals play a more significant role within a broader debate. The exploitation of tortoises in the Mediterranean Basin has been examined through the lens of optimal foraging theory and diet breadth models (e.g. [5-10]). According to the diet breadth model, resources are incorporated into diets based on their ranking and influenced by factors such as net return, which in turn depends on caloric value and search/handling costs [11]. Within these theoretical frameworks, tortoises hold a significant position. Their small size and sluggish movement require minimal effort and relatively simple technology for procurement and processing. This aligns with optimal foraging models in which the low handling costs of slow-moving prey compensate for their small size [5-6,9]. Tortoises also offer distinct advantages. They can be easily transported and kept alive, thereby maintaining freshness for deferred consumption [12-14]. For example, historical accounts suggest that Mexican traders recognised tortoises as portable and storable sources of protein and water [15]. Furthermore, tortoises provide non-edible resources, such as shells, which can serve as containers. This possibility has been discussed in the context of Kebara Cave [16] and noted in ethnographic and historical records (e.g. [12]). However, despite these advantages, their slow growth rate might have rendered intensive long-term predation unsustainable.
While tortoises are well-documented in the Southeast Asian archaeological record, zooarchaeological analyses in this region have been limited, particularly concerning prehistoric hunter-gatherer populations that may have relied extensively on inland chelonian taxa. With the present paper Bochaton et al. [17] aim to bridge this gap by conducting an exhaustive zooarchaeological analysis of turtle bone specimens from four Hoabinhian hunter-gatherer archaeological assemblages in Thailand and Cambodia. These assemblages span from the Late Pleistocene to the first half of the Holocene. The authors focus on bones attributed to the yellow-headed tortoise (Indotestudo elongata), which is the most prevalent taxon in the assemblages. The research include osteometric equations to estimate carapace size and explore population structures across various sites. The objective is to uncover human tortoise exploitation strategies in the region, and the results reveal consistent subsistence behaviours across diverse locations, even amidst varying environmental conditions. These final proposals suggest the possibility of cultural similarities across different periods and regions in continental Southeast Asia.
In summary, this paper [17] represents a significant advancement in the realm of zooarchaeological investigations of small prey within prehistoric communities in the region. While certain approaches and issues may require further refinement, they serve as a comprehensive and commendable foundation for assessing human hunting adaptations.
References
[1] Hartman, G., 2004. Long-term continuity of a freshwater turtle (Mauremys caspica rivulata) population in the northern Jordan Valley and its paleoenvironmental implications. In: Goren-Inbar, N., Speth, J.D. (Eds.), Human Paleoecology in the Levantine Corridor. Oxbow Books, Oxford, pp. 61-74. https://doi.org/10.2307/j.ctvh1dtct.11
[2] Alperson-Afil, N., Sharon, G., Kislev, M., Melamed, Y., Zohar, I., Ashkenazi, R., Biton, R., Werker, E., Hartman, G., Feibel, C., Goren-Inbar, N., 2009. Spatial organization of hominin activities at Gesher Benot Ya'aqov, Israel. Science 326, 1677-1680. https://doi.org/10.1126/science.1180695
[3] Archer, W., Braun, D.R., Harris, J.W., McCoy, J.T., Richmond, B.G., 2014. Early Pleistocene aquatic resource use in the Turkana Basin. J. Hum. Evol. 77, 74-87. https://doi.org/10.1016/j.jhevol.2014.02.012
[4] Blasco, R., Blain, H.A., Rosell, J., Carlos, D.J., Huguet, R., Rodríguez, J., Arsuaga, J.L., Bermúdez de Castro, J.M., Carbonell, E., 2011. Earliest evidence for human consumption of tortoises in the European Early Pleistocene from Sima del Elefante, Sierra de Atapuerca, Spain. J. Hum. Evol. 11, 265-282. https://doi.org/10.1016/j.jhevol.2011.06.002
[5] Stiner, M.C., Munro, N., Surovell, T.A., Tchernov, E., Bar-Yosef, O., 1999. Palaeolithic growth pulses evidenced by small animal exploitation. Science 283, 190-194. https://doi.org/10.1126/science.283.5399.190
[6] Stiner, M.C., Munro, N.D., Surovell, T.A., 2000. The tortoise and the hare: small-game use, the Broad-Spectrum Revolution, and paleolithic demography. Curr. Anthropol. 41, 39-73. https://doi.org/10.1086/300102
[7] Stiner, M.C., 2001. Thirty years on the “Broad Spectrum Revolution” and paleolithic demography. Proc. Natl. Acad. Sci. U. S. A. 98 (13), 6993-6996. https://doi.org/10.1073/pnas.121176198
[8] Stiner, M.C., 2005. The Faunas of Hayonim Cave (Israel): a 200,000-Year Record of Paleolithic Diet. Demography and Society. American School of Prehistoric Research, Bulletin 48. Peabody Museum Press, Harvard University, Cambridge.
[9] Stiner, M.C., Munro, N.D., 2002. Approaches to prehistoric diet breadth, demography, and prey ranking systems in time and space. J. Archaeol. Method Theory 9, 181-214. https://doi.org/10.1023/A:1016530308865
[10] Blasco, R., Cochard, D., Colonese, A.C., Laroulandie, V., Meier, J., Morin, E., Rufà, A., Tassoni, L., Thompson, J.C. 2022. Small animal use by Neanderthals. In Romagnoli, F., Rivals, F., Benazzi, S. (eds.), Updating Neanderthals: Understanding Behavioral Complexity in the Late Middle Palaeolithic. Elsevier Academic Press, pp. 123-143. ISBN 978-0-12-821428-2. https://doi.org/10.1016/C2019-0-03240-2
[11] Winterhalder, B., Smith, E.A., 2000. Analyzing adaptive strategies: human behavioural ecology at twenty-five. Evol. Anthropol. 9, 51-72. https://doi.org/10.1002/(sici)1520-6505(2000)9:2%3C51::aid-evan1%3E3.0.co;2-7
[12] Schneider, J.S., Everson, G.D., 1989. The Desert Tortoise (Xerobates agassizii) in the Prehistory of the Southwestern Great Basin and Adjacent areas. J. Calif. Gt. Basin Anthropol. 11, 175-202. http://www.jstor.org/stable/27825383
[13] Thompson, J.C., Henshilwood, C.S., 2014b. Nutritional values of tortoises relative to ungulates from the Middle Stone Age levels at Blombos Cave, South Africa: implications for foraging and social behaviour. J. Hum. Evol. 67, 33-47. https://doi.org/10.1016/j.jhevol.2013.09.010
[14] Blasco, R., Rosell, J., Smith, K.T., Maul, L.Ch., Sañudo, P., Barkai, R., Gopher, A. 2016. Tortoises as a Dietary Supplement: a view from the Middle Pleistocene site of Qesem Cave, Israel. Quat Sci Rev 133, 165-182. https://doi.org/10.1016/j.quascirev.2015.12.006
[15] Pepper, C., 1963. The truth about the tortoise. Desert Mag. 26, 10-11.
[16] Speth, J.D., Tchernov, E., 2002. Middle Paleolithic tortoise use at Kebara Cave (Israel). J. Archaeol. Sci. 29, 471-483. https://doi.org/10.1006/jasc.2001.0740
[17] Bochaton, C., Chantasri, S., Maneechote, M., Claude, J., Griggo, C., Naksri, W., Forestier, H., Sophady, H., Auertrakulvit, P., Bowonsachoti, J. and Zeitoun, V. (2023) Zooarchaeological investigation of the Hoabinhian exploitation of reptiles and amphibians in Thailand and Cambodia with a focus on the Yellow-headed Tortoise (Indotestudo elongata (Blyth, 1854)), BioRXiv, 2023.04.27.538552 , ver. 3 peer-reviewed and recommended by Peer Community in Archaeology. https://doi.org/10.1101/2023.04.27.538552v3

Faunal remains from the Upper Paleolithic site of Nahal Rahaf 2 in the southern Judean Desert, Israel
New zooarchaeological data from the Upper Palaeolithic site of Nahal Rahaf 2, Israel
Recommended by Ruth Blasco based on reviews by Ana Belén Galán and Joana GabucioThe Levantine Corridor is considered a crossing point to Eurasia and one of the main areas for detecting population flows (and their associated cultural and economic changes) during the Pleistocene. This area could have been closed during the most arid periods, giving rise to processes of population isolation between Africa and Eurasia and intermittent contact between Eurasian human communities [1,2].
Zooarchaeological studies of the early Upper Palaeolithic assemblages constitute an important source of knowledge about human subsistence, making them central to the debate on modern behaviour. The Early Upper Palaeolithic sequence in the Levant includes two cultural entities – the Early Ahmarian and the Levantine Aurignacian. This latter is dated to 39-33 ka and is considered a local adaptation of the European Aurignacian techno-complex. In this work, the authors present a zooarchaeological study of the Nahal Rahaf 2 (ca. 35 ka) archaeological site in the southern Judean Desert in Israel [3].
Zooarchaeological data from the early Upper Paleolithic desert regions of the southern Levant are not common due to preservation problems of non-lithic finds. In the case of Nahal Rahaf 2, recent excavation seasons brought to light a stratigraphical sequence composed of very well-preserved archaeological surfaces attributed to the 'Arkov-Divshon' cultural entity, which is associated with the Levantine Aurignacian.
This study shows age-specific caprine (Capra cf. Capra ibex) hunting on prime adults and a generalized procurement of gazelles (Gazella cf. Gazella gazella), which seem to have been selectively transported to the site and processed for within-bone nutrients. An interesting point to note is that the proportion of goats increases along the stratigraphic sequence, which suggests to the authors a specialization in the economy over time that is inversely related to the occupational intensity of use of the site.
It is also noteworthy that the materials represent a large sample compared to previous studies from the Upper Paleolithic of the Judean Desert and Negev.
In summary, this manuscript contributes significantly to the study of both the palaeoenvironment and human subsistence strategies in the Upper Palaeolithic and provides another important reference point for evaluating human hunting adaptations in the arid regions of the southern Levant.
References
[1] Bermúdez de Castro, J.-L., Martinon-Torres, M. (2013). A new model for the evolution of the human pleistocene populations of Europe. Quaternary Int. 295, 102-112. https://doi.org/10.1016/j.quaint.2012.02.036
[2] Bar-Yosef, O., Belfer-Cohen, A. (2010). The Levantine Upper Palaeolithic and Epipalaeolithic. In Garcea, E.A.A. (Ed), South-Eastern Mediterranean Peoples Between 130,000 and 10,000 Years Ago. Oxbow Books, pp. 144-167.
[3] Marom, N., Gnezdilov, D. L., Shafir, R., Barzilai, O. and Shemer, M. (2022). Faunal remains from the Upper Paleolithic site of Nahal Rahaf 2 in the southern Judean Desert, Israel. BioRxiv, 2022.05.17.492258, ver. 4 peer-reviewed and recommended by Peer community in Archaeology. https://www.biorxiv.org/content/10.1101/2022.05.17.492258v4

Wood technology: a Glossary and Code for analysis of archaeological wood from stone tool cultures
Open glossary for wood technologies
Recommended by Ruth Blasco based on reviews by Paloma Vidal-Matutano, Oriol López-Bultó, Eva Francesca Martellotta and Laura Caruso FerméWood is a widely available and versatile material, so it is not surprising that it has been a key resource throughout human history. However, it is more vulnerable to decomposition than other materials, and its direct use is only rarely recorded in prehistoric sites. Despite this, there are exceptions (e.g., [1-5] [6] and references therein), and indirect evidence of its use has been attested through use-wear analyses, residue analyses (e.g., [7]) and imprints on the ground (e.g., [8]). One interesting finding of note is that the technology required to make, for example, wooden spears was quite complex [9], leading some authors to propose that this type of tool production represented a cognitive leap for Pleistocene hominids [10]. Other researchers, however, have proposed that the production process for wooden tools could have been much easier than is currently thought [11]. Be that as it may, in recent years researchers have begun to approach wood remains systematically, developing analyses of natural and anthropogenic damage, often with the help of experimental reference samples.
In this work, the authors elaborate a comprehensive glossary as a first step towards the understanding of the use of wood for technological purposes in different times and places, as there is still a general gap in the established nomenclature. Thus, this glossary is a synthesis and standardisation of analytical terms for early wood technologies that includes clear definitions and descriptions of traces from stone tool-using cultures, to avoid confusion in ongoing and future studies of wood tools. For this, the authors have carried out a detailed search of the current literature to select appropriate terms associated with additional readings that provide a wide, state-of-the-art description of the field of wood technology.
An interesting point is that the glossary has been organised within a chaîne opératoire framework divided into categories including general terms and natural traces, and then complemented by an appendix of images. It is important to define the natural traces –understanding these as alterations caused by natural processes–because they can mask those modifications produced by other agents affecting both unmodified and modified wood before, during or after its human use.
In short, the work carried out by Milks et al. [6] is an excellent and complete assessment and vital to the technological approach to wooden artifacts from archaeological contexts and establishing a common point for a standardised nomenclature. One of its particular strengths is that the glossary is a preprint that will remain open during the coming years, so that other researchers can continue to make suggestions and refinements to improve the definitions, terms and citations within it.
[1] Oakley, K., Andrews, P., Keeley, L., Clark, J. (1977). A reappraisal of the Clacton spearpoint. Proceedings of the Prehistoric Society 43, 13-30. https://doi.org/10.1017/S0079497X00010343
[2] Thieme, H. (1997). Lower Palaeolithic hunting spears from Germany. Nature 385, 807-810. https://doi.org/10.1038/385807a0
[3] Schoch, W.H., Bigga, G., Böhner, U., Richter, P., Terberger, T. (2015). New insights on the wooden weapons from the Paleolithic site of Schöningen. Journal of Human Evolution 89, 214-225. https://doi.org/10.1016/j.jhevol.2015.08.004
[4] Aranguren, B., Revedin, A., Amico, N., Cavulli, F., Giachi, G., Grimaldi, S. et al. (2018). Wooden tools and fire technology in the early Neanderthal site of Poggetti Vecchi (Italy). Proceedings of the National Academy of Sciences. 115, 2054-2059. https://doi.org/10.1073/pnas.1716068115
[5] Rios-Garaizar, J., López-Bultó, O., Iriarte, E., Pérez-Garrido, C., Piqué, R., Aranburu, A., et al. (2018). A Middle Palaeolithic wooden digging stick from Aranbaltza III, Spain. PLoS ONE 13(3): e0195044. https://doi.org/10.1371/journal.pone.0195044
[6] Milks, A. G., Lehmann, J., Böhner, U., Leder, D., Koddenberg, T., Sietz, M., Vogel, M., Terberger, T. (2022). Wood technology: a Glossary and Code for analysis of archaeological wood from stone tool cultures. Peer-reviewed and recommended by PCI Archaeology https://doi.org/10.31219/osf.io/x8m4j
[7] Nugent, S. (2006). Applying use-wear and residue analyses to digging sticks. Mem Qld Mus Cult Herit Ser 4, 89-105. https://search.informit.org/doi/10.3316/informit.890092331962439
[8] Allué, E., Cabanes, D., Solé, A., Sala, R. (2012). Hearth Functioning and Forest Resource Exploitation Based on the Archeobotanical Assemblage from Level J, in: i Roura E. (Ed.), High Resolution Archaeology and Neanderthal Behavior: Time and Space in Level J of Abric Romaní (Capellades, Spain). Springer Netherlands, Dordrecht, pp. 373-385. https://doi.org/10.1007/978-94-007-3922-2_9
[9] Ennos, A.R., Chan, T.L. (2016). "Fire hardening" spear wood does slightly harden it, but makes it much weaker and more brittle. Biology Letters 12. https://doi.org/10.1098/rsbl.2016.0174
[10] Haidle, M.N. (2009). How to think a simple spear?, in: de Beaune S.A., Coolidge F.L., Wynn T. (Eds.), Cognitive Archaeology and Human Evolution. Cambridge University Press, New York, pp. 57-73.
[11] Garofoli, D. (2015). A Radical Embodied Approach to Lower Palaeolithic Spear-making. Journal of Mind and Behavior 36, 1-26.